In 2008 the National Institutes of Health launched the Human Microbiome Project with the aim of establishing a comprehensive catalog of microbial organisms living in or on the human body.1 Target domains included the gut, the skin, the urogenital tract, the oral and nasal mucosa and the ocular surface. A goal of this undertaking was to capitalize on the techniques of high-speed, high-fidelity nucleic acid sequencing developed for the human genome project to fully characterize the amalgam of commensal and pathogenic species that share our bodies with us.
After collecting 2.3 terabytes of sequence data in 35 billion sequencing reads, a picture of the immense diversity of microbes found in this search is beginning to emerge.2 For example, a 2011 study estimated that more than 200 species of bacteria from 59 genera commonly inhabit the human conjunctival mucosa.3 These species include an estimated 30 percent that represent novel or unclassified species.
The implications of this diversity are already finding their way into the clinic in other areas of medicine, where particular microbiota have been associated with conditions including obesity, asthma, psoriasis and a number of gastrointestinal disorders.2 This month, we’ll consider the microbiome of the ocular surface, examine how microbial species interact and coexist within this unique environment, and then explore how advances in our understanding of microbial dynamics may impact ocular therapeutics going forward.
Microbiome Building Blocks
Bacteria from the environment and from surrounding tissues colonize the conjunctival and corneal surface, as well as associated tissues (lacrimal and meibomian ducts) early in life. Typical residents found in low numbers include gram-positive species such as Staphylococci, Corynebacterium, Streptococcus and Propionibacterium.3,4 Gram-negative species such as Neisseria or Pseudomonas are less common, as are typical pathogens such as S. aureus or H. influenza. Regardless of the specific inhabitants, this bacterial melting pot establishes a communal equilibrium that can remain stable indefinitely.
It’s thought that in the presence of an intact epithelial barrier, commensal species act in several ways to protect the ocular surface from pathogenic infection.4 First, the innocuous commensal species serve as “space holders,” preventing more virulent strains from gaining a foothold in the ocular environment. Second, a low level of activation of the innate immune system is primed by resident species and keeps the ocular surface at the ready should more pathogenic species invade. Other studies have shown an active interaction between bacteria and the mucins of the ocular surface, a complex relationship that may have both commensal and pathogenic implications.5,6
The mucins of the ocular surface exist in both soluble and membrane-associated forms, and one of their key functions is to act as anti-adhesives, preventing bacteria from establishing a foothold. Soluble mucins also have antimicrobial properties that contribute to the tear film’s bacteriostatic role.7 In addition, some bacteria produce enzymes that cleave membrane-associated mucins from the epithelial surface.7 It’s possible that this process is part of the ecosystem of the ocular surface: Both mucins and their bacterial guests have been coevolving for millions of years.
| ||||||||||||||||
Investigating Bacteria
When it comes to our understanding of the world of microorganisms, it’s hard to underestimate the biological and clinical value of the bacteria streaked on an agar plate, or the yeast cultured in a flask of broth. The ability to isolate and culture microbes was unequivocally a paradigm-shifting event in the history of medicine. And yet there are few fields in medicine that are so wed to their experimental constructs as microbiology. In its earliest days, the isolated cultures were the only way the microbiologist could identify, characterize and select appropriate treatments for disease causing pathogens.
The methods of classical microbiology later made possible the tremendous advances of 20th-century biomedicine, from decoding of DNA to the fundamentals of gene expression and repression. Along the way, however, a critical concept was overlooked: The laboratory is not the natural environment for bacteria or other microbial organisms. Perhaps more important is the corollary to this concept: Just because a bacterial strain won’t grow on a Petri dish doesn’t mean it won’t still grow well on the surface of your eye.
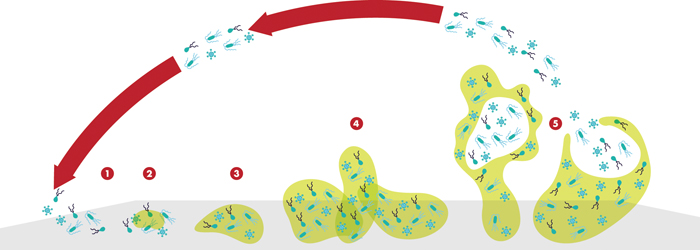
Although the free-floating, planktonic form of most bacterial species is the one that we are most familiar with from microbiology labs, the life cycle of most bacteria is dominated by a sessile phenotype, embedded in a hydrated matrix of secreted polymers. In their native habitat, microbes most often live in ultra-thin layers coating any and all available surfaces, from catheters to contact lenses. As they establish new habitats their secretions construct a complex film-like meshwork that provides additional surface area for bacterial expansion, and as it turns out, another layer of defense against antimicrobial control. These coatings, whether it’s the slime of a river cobble or the plaque on your teeth, are all examples of biofilms, a term coined by the Canadian microbiologist J.W. (Bill) Costerton in the early 1980s.8 Bacterial biofilms form wherever there is water and air—and sometimes they don’t even need the air. The complexity and variety of these structures is hard to underestimate, as is their importance in disparate fields including medicine, manufacturing and waste management.
Biofilms and the Ocular Space
The ocular surface provides a less-than-ideal setting for biofilm formation, with the constant physical disruption of blinking and tear flow. In addition, the tear film contains antibacterial components (in addition to mucins), including lactoferrin, lysozyme and defensins, that hamper microbial growth.4 Under these conditions, macroscopic biofilms are unlikely to form, but smaller regions of bacterial association are still possible. Biofilms become a bigger issue when abiotic surfaces such as contact lenses, intraocular lenses and punctal plugs are introduced into the ocular surface.4 Any of these artificial surfaces can provide a starting point for biofilm development.
|
A simpler approach used in several other recent studies compared the profile of species from asymptomatic controls with those from patients with keratitis or blepharitis.11,12 One study found Achromobacter and Stenotrophomonas were prevalent in all keratitis samples from contact lenses (n=28), but were present in only one asymptomatic individual (n=9).11 A second study collected bacteria from tears and eyelashes of patients with blepharitis, and compared the species found with those on control samples.12 While the distribution of species was diverse, there was a clear, significant pattern of genera present in patients who were symptomatic that was not seen in controls.
These screening approaches do not directly establish a role for biofilm-related genes or the disease-associated species in ocular infection pathogenesis, but they do highlight the idea that changes in the phenotype and abundance of bacteria are important in the disease process.13
We have a lot to learn about the microbial ecology of the ocular surface. Are we close to applying this knowledge to new therapies? One aspect of biofilm ecology that may lead to treatments is the quorum-sensing systems used by all bacteria to communicate within the biofilm.14 It’s thought that one of these signaling pathways may allow us to alter the properties of bacteria, making them more susceptible to traditional antibiotic therapies. Beyond this, we should keep our eyes on developments in other aspects of the human microbiome, as we begin to see parallels in the interaction between our bacterial guests and the surfaces they inhabit. Perhaps we may eventually develop a probiotic for ocular therapy. REVIEW
Dr. Abelson is a clinical professor of ophthalmology at Harvard Medical School and senior clinical scientist at the Schepens Eye Research Institute. Dr. McLaughlin is a medical writer at Ora in Andover.
1. Turnbaugh P, Ley RE, Hamady M, Fraser-Liggett CM, Knight R, Gordon JI. The Human Microbiome Project. Nature 2007;449:804-810.
2. Cho I, Blaser MJ. The human microbiome: At the interface of health and disease. Nature Rev. Genetics 2012;13:260-270.
3. Dong Q, Brulc JM, Iovieno A, et al. Diversity of bacteria at healthy human conjunctiva. Invest Ophthalmol Vis Sci 2011;52:5408–5413.
4. Miller D, Iovieno A. The role of microbial flora on the ocular surface. Curr Opin in Allergy Clin Immunol 2009;9:466-470.
5. Mantelli F, Argueso P. Function of ocular mucins in health and disease. Curr Opin Allergy Clin Immunol 2008;8:477–483.
6. Berry M, Corfield AP, Harris A, Khan-Lim D. Functional processing of ocular mucins. Adv Exp Med Biol 2002;506:283–288.
7. Berry M, Harris A, Lumb R, and Powell K. Commensal ocular bacteria degrade mucin. Brit J Ophthal 2002;86:1412-1416.
8. Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial Biofilms: From the natural environment to infectious diseases. Nature Rev. Microbiology 2004;2:95-108.
9. Duggirala A, Kenchappa P, Sharma S, et al. High-resolution genome profiling differentiated Staphylococcus epidermidis isolated from patients with ocular infections and normal individuals. Invest Ophthalmol Vis Sci. 2007;48:3239-45.
10. Wenbo Hou W, Sun X, Wang Z, Zhang Y. Biofilm-forming capacity of staphylococcus epidermidis, staphylococcus aureus, and pseudomonas aeruginosa from ocular infections. Invest Ophthalmol Vis Sci 2012;53:5624-5631.
11. Wiley L, Bridge DR, Wiley LA, Odom JV, Elliot T, Olson JC, Bacterial biofilm diversity in contact-lens related disease: Emerging roles of Achromobacter, Stenotrophomonas, and Delpha. Invest Ophthalmol Vis Sci 2012;53:3896-3905.
12. Se Hee Lee SH, Oh DH, Jung JY, Kim JC, Jeon CO. Comparative ocular microbial communities in humans with and without blepharitis. Invest Ophthalmol Vis Sci 2012;53:5585-5593.
13. Zegans ME, Shanks RMQ, O’Toole GM. Bacterial biofilms and ocular infections. Ocular Surface 2005;3:73-80.
14. Njoroge J, Sperandio V. Jamming bacterial communication: New approaches for the treatment of infectious disease. EMBO Mol Med 2009;1:201-210.